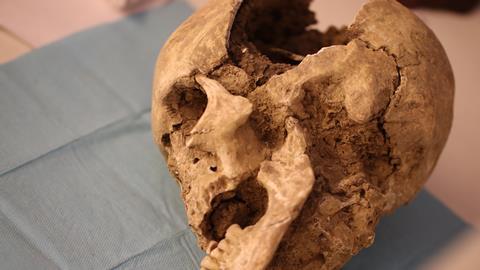

Harbouring probably more than 700 species, the oral microbial community is retained to be the second most biodiverse ecosystem after the gut. However, unlike the gut, the extent of most of the variables that affect the oral microbiome composition remain less known.
Despite being quite resilient, we know that therapies (e.g. antibiotics), diseases, hygienic practices as well as the interaction with the host, are able to shape the overall microbial composition. The oral geography itself can also affect the presence of microorganisms in specific oral niches. The influence of diet is more questionable depending on the oral niche we are considering, even if some evidence in this regard are now being reported in the scientific literature. Diet has always played a major role in the evolutionary story of our species, directly influencing our genome. However, how human dietary choice might have influenced the evolution of our symbionts remain largely unanswered.
Therefore, we aimed to investigate whether such a revolutionary change in our history as the introduction of agriculture (~10,000 BCE) could have left some marks on oral microbial community composition. Exploring key passages in human history from a microbial perspective, can reinstitute crucial information related to both microbial taxa evolution and, possibly, to the evolution of our life condition. We collected dental calculus samples, which have been proven to preserve ancient bacterial and human biomolecules in such a quality that allow the reconstruction of entire ancient bacterial genomes. The selected samples (i.e. 76) belonged to different Neolithic (6,200-4,000 BCE), Post-Neolithic (3,500-2,000 BCE) and Pre-Neolithic (Paleolithic, 31,000-11,000 BCE) small communities, mostly from Apulia region (Southern Italy), in the same areas that saw the transition to agriculture in the seventh century BCE.
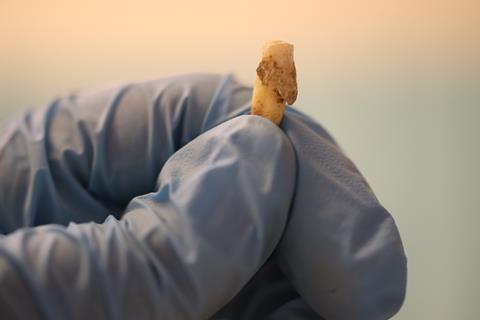
Having several coeval and diachronic communities from the same geographical area at our disposal, we were able to detect the changes, in terms of microbial composition, both between contemporary communities and with respect to previous and subsequent periods of transition.
Our study revealed that the oral microbiome underwent two different shifts during the considered period – probably linked to the changes in subsistence strategies. After a long phase in the upper Paleolithic in which the composition of the microbiome of the hunter-gatherers remains stable, we detected a first change in the early centuries of the transition to agriculture. The hunter-gatherers’ microbiome was composed by an important presence of Actinomyces spp., Streptococcus HOT 061, Streptococcus sanguinis, and a low abundance of other symbionts, such as all the species belonging to the so called red and orange complexes. With the beginning of the transition, the abundance of distinct species which are known oral pathobionts, such as Porphyromonas gingivalis, Tannerella forsythia and Treponema denticola start to rise, along with other species quite unfrequently found nowadays, such as Olsenella sp. oral taxon 807. In the last centuries of the Neolithic period, we detected a second shift in the oral composition, where these characteristics became more accentuated, and the abundance of these species became more prominent. At the same time the taxonomic characteristics of the earlier hunter-gatherer samples becomes less abundant.
Knowing the influence that diet can exert on human symbiont communities, and that micro-debris of ingested food can be found directly within the dental calculus itself, we integrated our metagenomic analysis with the microscopic identification of food remains embedded in the dental calculus of the same subjects. We aimed to find possible correspondence between the metagenomic profile and the dietary information. Moreover, some of the considered populations had already been studied for their dietary intake using isotope analyses, thus giving further information about their subsistence strategies. Within the Palaeolithic samples, a great number of vegetal elements has been found entrapped in the dental calculus, in particular starch granules. This finding provided an interesting correspondence with the bacterial functional profile, where indeed, an enrichment in starch degrading pathways was found in pre-Neolithic samples, compared to the Neolithic and post-Neolithic ones.
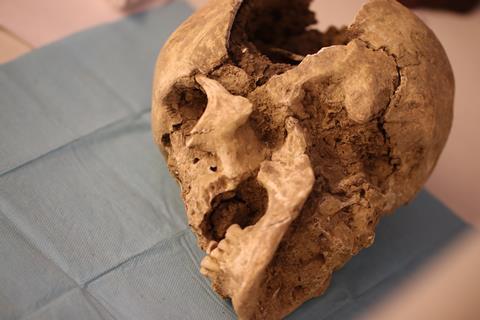
Other anthropological analysis on this population reported that in addition to the consumption of proteins and animal fats, vegetable intake constituted a huge part of the hunter-gatherer communities’ diet. Neolithic samples, on the other hand, showed a lower variability of plant species, and the oral bacteria of the Neolithic and post-Neolithic communities have a different functional profile, enriched in species that metabolize galactose (a very abundant sugar in dairy products) as well as glycan and polysaccharides. One last interesting aspect was the higher presence of virulence factor pathways among all the Neolithic samples, especially in the second phase of transition at the end of the Neolithic period. Even in this case, the higher incidence of pathobiont species and virulence pathways were in accordance with the higher incidence of oral pathologies founded among samples belonging to the last centuries of the Neolithic period, and also to the following period considered, as reported by the anthropological analysis of skeletal remains.
Overall, the taxonomic and functional profile of Neolithic microbial communities showed many points of overlap with the oral microbiota (i.e. from saliva and dental plaque) of present-day individuals with a very consistent (or even exclusive) vegetable and carbohydrate intake in their diet, thus suggesting a diet particularly rich in these elements. For example, species as Capnocytophaga spp., Ottowia HOT 894, Neisseria elongata and Rothia aerea, that were distinctive of the agricultural populations, have recently been associated with a higher consumption of dietary fibre from dental plaque. Also the enrichment in glycan degradation pathways has been observed in chimpanzees, characterized by an exclusive plant-based feeding subsistence.
The hypothesis that the observed change in oral bacterial profile could be associated with diet was corroborated by isotope analysis conducted on these populations, showing an increase in vegetable intake (especially cereals and legumes) from the last part of the Neolithic period, and a very poor intake in animal proteins. Combining the metagenomic profile with the obtained botanical information and isotope data, we can probably hypothesize that the studied agricultural societies based their diet on a much less diverse resource network, especially at the end of the Neolithic period, while the first centuries seem to be halfway between the established agricultural lifestyle and the pre-agricultural one. This reduction in terms of the variability of consumed plants and a very poor intake of animal protein, might have favoured the presence of specific bacterial taxa which thus became prominent in the oral microbiome community.

Other hypotheses should be considered to possibly explain some of the observed changes in the aforementioned species abundance, such as the influence of bacterial genome evolution. Indeed, by reconstructing six ancient genomes of Olsenella sp. oral taxon 807, one of the most interesting biomarkers associated with the transition to agriculture, we found that several regions involved in defence mechanisms and interactions with other microbial commensals (e.g. CRISPR genes) were missing. These traits are expected from an evolutionary perspective, since competition is a major force influencing bacterial evolution, especially in that ecological niche where strong interactions among different species and metabolic overlap is expected. The development of defence mechanisms can deeply affect species fitness and also change the network of relations among species within a same ecological niche. Our data on Olsenella sp. oral taxon 807, suggests that its ability to compete and interact with other oral species living in the same niche has evolved recently.
The results obtained from the analysis of ancient Neolithic oral microbiomes seemed to suggest that the bacterial composition had gradually and continuously adapted to the new system of agricultural subsistence, veritably not improving the oral health condition of the studied population.






No comments yet