
Imagine an environment so extreme that most life cannot survive: a pitch-dark cave deep beneath the mountains of Northern Spain, or a hyper-arid desert in Chile where rainfall is virtually non-existent. These are not lifeless wastelands. Beneath cave walls and within the dusty top layer of desert soils, thriving communities of cyanobacteria, green algae, and fungi quietly engineer their ecosystems: fixing carbon, weathering rock, and cycling nutrients in conditions that would defeat most organisms on Earth.
Our research group XCEL – Extreme Cryptogam Ecology Lab at the University of Applied Sciences Kaiserslautern in Germany, has spent years studying these cryptic communities. In two recent studies, we used classical culture-dependent approaches to isolate and characterise the microbiomes of a natural cave in Northern Spain and the grit crust of the Atacama Desert in Chile. Together, these investigations reveal a remarkable breadth of microbial diversity, including potentially new genera and species, and underscore why putting these microbes “in bottles” remains one of the most powerful tools in microbiology.
Why culture?
In the age of metagenomics and high-throughput sequencing, the humble agar plate might seem outdated. Metabarcoding can catalogue thousands of taxa from a single environmental sample, but it comes with a catch: it recovers DNA from living and dead organisms alike, and the short sequence fragments it produces often lack the resolution needed to distinguish closely related species.
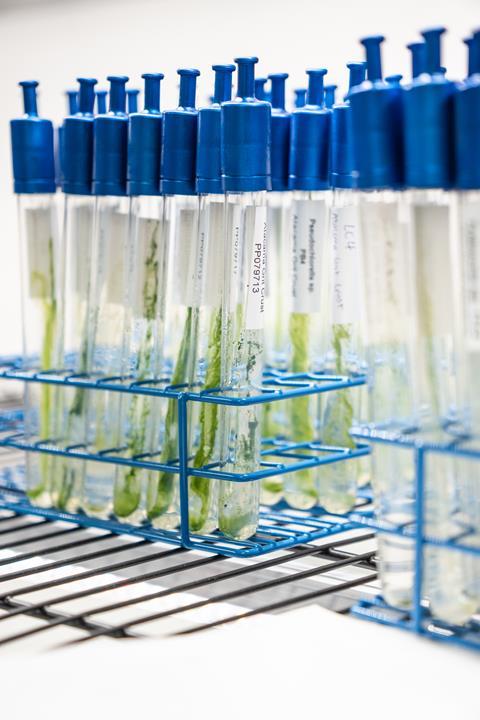
Culturing microbes - isolating them on selective media, growing them under controlled conditions, and studying them one by one - offers something sequencing alone cannot. It confirms that an organism is alive and metabolically active, it yields high-quality DNA for robust phylogenetic analysis. And, crucially, it produces living reference strains that can be stored in culture collections, shared between laboratories, and used in future experiments, from genome sequencing to biotechnological screening. In our studies, cultural methods even recovered rare taxa representing less than 1% of amplicon reads in earlier metabarcoding surveys. Such organisms might otherwise have been dismissed as noise in a dataset.
A glimpse into darkness: life in a cave
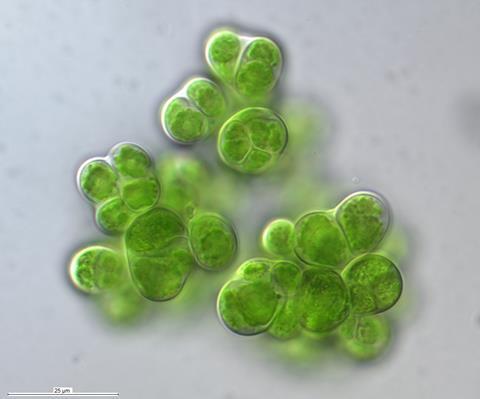
Caves are often imagined as barren, but those with even the faintest traces of natural light harbour surprisingly complex communities. Subaerial biofilms known as thin, slimy crusts that coat cave walls, are frequently dominated by cyanobacteria, the ancient photosynthesisers that oxygenated Earth’s early atmosphere over two billion years ago. Accompanied by green algae, fungi and sometimes bryophytes, these biofilms form multi-layered, three-dimensional micro-ecosystems where each organism occupies a specific niche.

We collected biofilm samples from a restricted natural cave near Bilbao and cultivated the organisms on three types of selective media, each targeting a different group: BG11 for cyanobacteria, Bold’s Basal Medium for green algae, and Sabouraud agar for fungi. Over two years of patient subculturing, we generated a total of 123 isolates: 58 cyanobacteria falling into 21 genera, 24 green algae in 10 genera, and 41 fungi in 13 genera.
Among the cyanobacteria, we encountered some truly remarkable organisms. Gloeobacter species, which probably evolved around 3.4 billion years ago, formed striking purple biofilms in the cave. Unlike almost all other cyanobacteria, Gloeobacter lacks internal thylakoid membranes - the structures where photosynthesis typically takes place - making it a living relic of early photosynthetic evolution. Alongside these ancient lineages, we isolated calcifying species such as Scytonema julianum, which produces tubular sheaths of calcium carbonate around their filaments. Even more impressive was a member of the genus Geitleria which produces even thicker calcified sheaths made of calcite crystals in a well-oriented manner. Similar to attempts made by colleagues reported for Geitelria appalachiana found in the Appalachian mountains, we were unable to isolate this cyanobacteria, which is bothering the whole team. These mineralised structures not only extend the biofilm into three-dimensional space, creating micro-habitats for other organisms, they also offer unique chances for future biotechnological applications.

The cave also yielded typical subterranean specialists: Chalicogloea cavernicola, Timaviella karstica with its distinctive purple filaments, and Oculatella subterranea, recognisable by the characteristic orange eyespot in its apical cell. Several isolates did not match any described species and likely represent new genera - a finding that supports the idea that caves act as isolated evolutionary niches, fostering niche specialisation among cyanobacteria.
The fungal community told a different story. Dominated by decomposers and phytoparasites such as Sporobolomyces, Stereum, and Phlebia, the fungi appeared to be largely trogloxenic: visitors that entered the cave from the surrounding mixed forest via wind, water, or animal activity. Enzyme assays revealed a broad suite of carbohydrate-degrading, lipid-hydrolysing, and phosphate-mobilising activities, confirming that these fungi are active participants in nutrient cycling within the biofilm. They decompose the organic matter produced by the phototrophs, effectively closing the loop in an underground carbon cycle.
The desert in bottles: life in the Atacama’s grit crust
If a Spanish cave is an unusual habitat for photosynthetic life, the coastal Atacama Desert pushes the concept to its absolute limit. Parts of the Atacama receive virtually no rainfall, and ground-level temperatures regularly exceed 50°C. Yet in 2020, our group described a previously unknown type of biological soil crust, the grit crust, forming on tiny granitoid pebbles in the National Park Pan de Azúcar. Here, lichens colonise translucent stones just 2–6 mm in diameter at such density that their dark pigments create a checkerboard pattern visible across the landscape.

While the lichens and their photobionts (symbiotic green algae of the genus Trebouxia) form the backbone of the grit crust, our latest study focused on the non-lichenised, free-living microorganisms accompanying them. Using the same integrative cultivation approach, we isolated 122 organisms from four sites arranged along a moisture gradient - from a relatively humid coastal ridge, frequently bathed in fog, to a barren inland location 12 km from the sea with almost no fog influence.
The results were striking. We recovered 38 cyanobacteria, 26 green algae, and 58 non-lichenised fungi. The community composition shifted clearly along the water availability gradient. At the humid, fog-influenced coastal site, green algae and lichens dominated, producing the highest soil carbon and nitrogen contents. At the driest inland sites, where fog rarely penetrated, cyanobacteria took over. Microclimate data helped explain this pattern: while many lichens and green algae can reactivate their photosynthesis from high humidity alone, cyanobacteria require liquid water, even if it is just a thin film of dew condensing on a pebble at night. At the driest site, the pebbles themselves appear to act as tiny condensers, harvesting enough moisture from temperature-driven condensation–evaporation cycles to sustain cyanobacterial life.
Most of the isolated cyanobacteria matched this ecology because they belonged to the order Chroococcidiopsidales, unicellular taxa long celebrated for their extreme stress tolerance and considered model organisms in astrobiology. However, none of our isolates matched any previously described genus within this order. Instead, they fell into several distinct, well-supported phylogenetic clades, each with its own morphological characteristics, representing potentially new genera unique to the grit crust of the Atacama Desert. Having these lineages in culture is a striking advancement since other members of the Chroococcidiopsidales live as symbionts in lichens and the relationship between free-living and symbiontic representatives has not been investigated, yet but can now be studied.
What makes these microbes special?
Beyond the sheer number of new taxa, what stands out is the functional coherence of these communities. In both the cave and the desert, the same basic ecological architecture repeats: cyanobacteria (and green algae) serve as primary producers, fixing carbon and, in the case of Nostoc species, atmospheric nitrogen. Their extracellular polymeric substances glue communities together, retain moisture, and protect against UV radiation. Fungi, in turn, decompose and recycle organic matter, mobilise phosphorus, and structurally reinforce the biofilm or biocrust with their hyphae. These are not random assemblages but tightly integrated micro-ecosystems with their own microbial architecture, shaped by millions of years of co-evolution under extreme selective pressures.
The cave and desert organisms also hold practical promise. Chromochloris zofingiensis, a green alga we isolated from the cave, is an emerging model for astaxanthin production, a high-value ketocarotenoid with antioxidant properties used in aquaculture and nutraceuticals. The cave isolate, adapted to extremely low light, may have altered pigment-production pathways compared to standard laboratory strains, making it a potential candidate for biotechnological exploitation. Similarly, the extremotolerant Chroococcidiopsidales from the Atacama are of interest to astrobiologists modelling the limits of life on Mars-like surfaces.
Culturing the unculturable
A persistent myth in microbiology is that most environmental microbes are “unculturable.” Our work challenges this narrative - not by denying that cultivation captures only a fraction of total diversity, but by showing that the fraction it does capture is irreplaceable. Every isolate is a living resource. It can be sequenced, experimentally characterised, and taxonomically described. It anchors the short, anonymous DNA fragments generated by metabarcoding to real, observable organisms with known morphologies and ecologies.
As, for example, cyanobacterial taxonomy undergoes a major overhaul (the phylum was recently restructured into 20 orders and 43 families) and curated databases such as CyanoSeq allow more accurate assignment of environmental sequences, the demand for well-characterised reference strains has never been greater. Culture collections are the bedrock on which modern microbial taxonomy, ecology, and genomics are built.
Looking ahead
Our culture collections from the Spanish cave and the Atacama grit crust now represent one of the most comprehensive sets of isolates from any biocrust or cave biofilm system worldwide. Future work will combine these living strains with whole-genome sequencing, population-genomics approaches, and targeted physiological experiments. We are particularly interested in understanding how the novel Chroococcidiopsidales genera from the Atacama differ from their relatives in other deserts or the ones living in lichen symbiosis, what molecular mechanisms enable calcification in cave cyanobacteria, and how fungi interact with their phototrophic partners at the biochemical level.
The next step is clear: to bridge the gap between culture-dependent and culture-independent methods, using isolates as anchors to interpret the vast, often anonymous, datasets generated by metagenomics. By doing so, we can move from cataloguing diversity to understanding it, and from understanding it to harnessing it, whether for biotechnology, conservation, or the fundamental science of how life persists at the extremes.
Further Reading
Jung, P., Briegel-Williams, L., Nürnberg, D.J., Wolf, T., Guillen, A., Leira, M. & Lakatos, M. (2025). A glimpse into darkness: Diversity of culturable cyanobacteria, green algae and fungi from subaerial cave biofilms. Journal of Phycology, 61, 1699–1717. https://doi.org/10.1111/jpy.70104
Jung, P., Briegel-Williams, L., Werner, L., Jost, E., Brand, R., Baumann, K. & Lakatos, M. (2025). The Desert in Bottles: The Culturable Microbiome of the Atacama Desert’s Grit Crust. Environmental Microbiology Reports, 17, e70194. https://doi.org/10.1111/1758-2229.70194
Funding Acknowledgements
This work was funded by the German Research Foundation (DFG; JU 3228/1-1) and the Carl-Zeiss Foundation (P2023-03-051). The author declares no conflicts of interest.





No comments yet